Genomi on jokaisessa valmiina, kun vain sille annetaan mahdollisuus myrkyttömästi toimia. Aivot ovat rasvamoduli, hyvin rikas kaikenlaisista monimutkaisista lipidirakenteista, siis hyvin kaunis rakenne! Jos ne kulutetaan orgaanisilla liuottimilla (joka alkoholi mielestäni on), ne eivät voi muodostua yhtä pian takaisin kuin mitä ne hajoavat siinä tapauksessa, varsinkin jos estävä toxinen aine jatkuvasti tarjoutuu aivojen rasvojen uudistuskoneistoon.
Normaalisti aivot uudistanevat rasvojaan sitä tahtia kuin Harper 1970- luvun taitteessa sanoi: 20% aivojen rasvoista uudistuu viikossa. Siis aivot uudistuvat joka päivä jonkin verran rasvoiltaan( ei varsinaisilta neuroneilta ja tukisoluilta , vaan niiden rakenteellisilta ja kalvoaineilta, joten jokaisen päivän raittius on merkitsevä asia. Ihmisellä on vain tämä päivä. Jos on tämän päivän raitis, on ollut raittiina yhden tärkeimmän ajan tulevaisuudesta, jota ei vielä ole, sillä tämä päivä on.
Itseasiassa viidessä vuodessa voi saada aivojensa rasvamodulin aika hyvän kuntoon, jos on ollut raittiina ja syönyt monipuolista ravintoa ja harrastanut ulkoilua ja liikuntaa, niin että lihasten ja luustonkin kunto on kohentunut, sillä tärkeälle kalsiumin tasapainolle on eduksi, että on liikuntaa ja luusto kunnossa.(Kaikkiin tärkeisiin reaktioihin arvitaan kalsiumjoneja oikeissa paikoissa oikealla hetkellä).
Alkoholin vaikuttama fosfatidyylietanoli-peräinen, vain alkoholista johtuva fosfolipidimuoto PEth, kroonisessa alkoholismissa ottaa tilaa aivolipideissä ja estää monipuolisen lipidikirjon tasapainoisen muodostumisen kehossa, ei vain aivoissa, vaan lopulta kaikkien tärkeiden elinten solukalvoissa.
Alkoholin myrkystä pitää normaalilla ravinnolla ja liikunnalla vieroittua ja antaa taustalla olevan soluaarteen, esi-isiltä perityn genomin, korjata soluja. Ihmisen keholla on ihmeellinen taipumus korjaantua, kun sitä ei myrkytetä millään toksiinilla. PEth moklekyylille ei ole mitään luonnollista hajottavaa entsyymiä kehossa valmiina, vaan se puoliintuu molekyylin fysikokemiallisten vanhenemislakien mukaan muutamissa päivissä. (Jokainen molekyyli "vanhenee" sidoksissaan ajallaan) .
Katselin netistä ihan tavallista tekstiä tavallisen fosfatidyylietanolaminin muodostuksesta.
Biosynthesis of phosphatidylethanolamine (PE).
http://lipidlibrary.aocs.org/Biochemistry/content.cfm?ItemNumber=39191
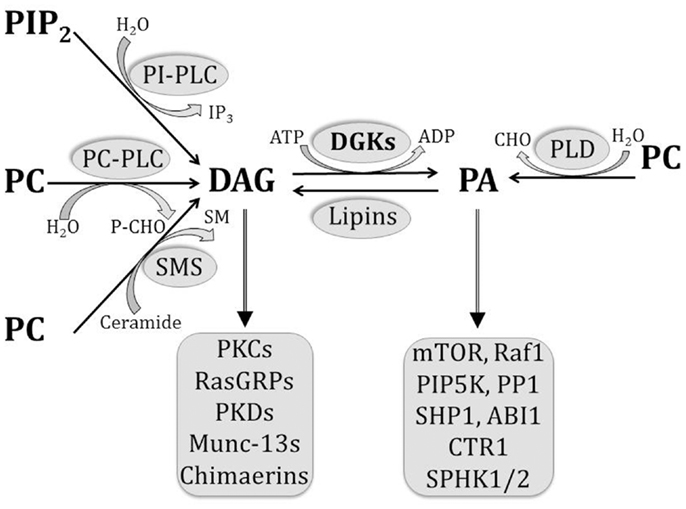
Sen merkki on PE.( Sitä sänotaan kefaliiniksi. Lesitiinin merkki on PC. Se on fosfatidyylikoliini. Fosfatidyyli-inositolin merkki on PI ja sita sanotaan lipositoliksi. Se voi rikastua fosfaateilla ja olla solukalvoissa muotoja PIP, PIP2, PIP3. Varsinkin aivojen harmaissa soluissa on tämä PI-energiajärjestelmä käytössä ja monessa muussakin spesifisessä prosessissa.
Lisäksi solukalvoissa on sfingomyeliiniä (SM).
Kalvot ovat kaksoislipidikerroksia ja nämä fosfolipidit (PL) rikastuvat rasvahapoilla, osin niistä, joita ihmiset syövät ja jotka muokkautuu kehossa tai joita muodostuu kehossa etikkahaposta ( tai propionihaposta) käsin energia-aineenvaihdunnan puolelta. .
Rasvahapot ovat lipidiliukoisia päädyltään ja niitä asettuu solun lipidikaksoiskalvoon rasvaliukoiset päädyt sisäänpäin kalvon keskiosiin. Täten kalvon sisäosa on hyvin hydrofobinen ja siellä on myös kolesterolimolekyylejä tiivistämässä kalvoa vedenpitäväksi.Keholle kolesteroli on tärkeä. keho tekee jokaiseen soluunsa kolesterolia vedenpitävyyden varmistamiseksi ja kehon endogeeninen kolesterolin tuotanto on monikertainen verrattuna siihen määrään, mikä pidetään ravinnossa saatavan kolesterolin suosituksen ylärajana ( 300 mg päiväss, yksi kananmunan keltuainen). Keho syntetisoi kolesterolia fundamentaalisesta aineenvaihdunnasta käsin. Nykyaikana vain jokin tekijä on häirinnyt tätä järjestelmää, ehkä ravinnon laatu ja energian ylitarjonta, joten joudutaan jarruttamaan endogeenista kolesterolin tuotantoa toisilla ja rajoittamaan exogeenistakin kolesterolipisaraa. Mutta esim eläimet luonnossa eivät näy omaavan kolesteroliongelmia. ja toisaalta - liekö kenenkään huoli, jos karjalla on korkeat kolesterolit.
Fosfolipideillä PE, PC, PS, PI, PS ja sfingomyeliini SM on tietty asemansa ( ja tehtävänsä) tässä plasmakalvossa. Ne sijoittuvat riippuen niiden hydrofobisuuden ja hydrofiilisyyden asteesta. Ne eivät sijaitse esim vierekkäin tai päällekkäin kuin seinätiilit, vaan kelluvat asymmetrisesti ominaisissa asemissaan. Lähimpänä solun sisäosan sytosolia on fosfatidylseriini PS. Se on eräänlainen signaali kalvorakenteeen dynaamisestä elinkyvystä, jossa energisesti saadaan pidettyä kukin fosfolipidilaji omalla paikallaan. Jos PS pulpahtaisi solun pintaan (kuin kuollut kala nurin niskoin), se toimisi merkkinä. "Olen vanhentuneesta solusta. Poimikaa minut pois joukosta#. Ja silloin Soluyhteisö poimii sen solun pois joukosta. Kts. kuvaa punasolun kalvosta! Punasolun ikä on 120 päivää, sitten se poimiutuu pois. Valmiissa punasolussa ei nimittäin ole koodia enää sisällä.
http://www.rbclab.com/Pages/500/520/JPEG/Figure%2015.1%20png%20copy.png
Paljon siitä ihmisen syömästä energiasta kuluu tähän kehon solukalvon dynaamisuuden ja polaarisuuden ylläpitoon, elämäntaisteluun solutasossa, siihen "virtaan", siis solujen energiatilan ylläpitoon, sillä elävässä solussa saadaan tämän kalvon avulla kehitettyä erilaisia potentiaalieroja .
Koko kehon koko soluyhteisön elämän voimien integroituna summana ihminen voi tuntea itsensä pirteäksi tai myöskin väsyneeksi.
Tietysti on myös neutraalit kalvopumput jotka katsovat että solut ovat neutraaleja.
Monet solutyypit ovat erikoistuneet toimimaan näiden aktiopotentiaalien avulla niin että voidaan mitata sähkökäyrää ( neuronit, EEG, betasolu oskillaatio, sydän EKG, Silmän retinogrammi) . Mutta jos on tällaisia erityisiä sähköisesti ärtyviä soluja, niiden ympärillä on isolaatiosoluja, jotka eivät ole sähköisesti ärtyviä. Näissä asioissa on solukalvot olennaisen tärkeitä. Ne antavat isoloivaa kaapelimateriaalia ja kantavat myös neutraloivia pumppuja. Jos kaikessa tässä kalvomateriaalissa on voittopuolisesti lipidikirjon sijasta vain jotakin fosfatidyyliPethmateriaalia- ei voi odottaa että henkilön terveydentunne olisi kehuttava. Voisi sanoa että pikemminkin pyton olo. Kaikista lääkkeistä mikään ei vedä vertoja raittiudella ja tavalliselle ruoalle.
Phospholipid flip-flop and scrambling
https://www.ncbi.nlm.nih.gov/pubmed/15809683
Cell Mol Biol Lett. 2005;10(1):101-21.
The mystery of phospholipid flip-flop in biogenic membranes.- Cell Mol Biol Lett. 2005;10(2):363.
Phospholipid
flip-flop is required for bilayer assembly and the maintenance of
biogenic (self-synthesizing) membranes such as the eukaryotic
endoplasmic reticulum and the bacterial cytoplasmic membrane. Due to the
membrane topology of phospholipid biosynthesis, newly synthesized
phospholipids are initially located in the cytoplasmic leaflet of
biogenic membranes and must be translocated to the exoplasmic leaflet to
give uniform bilayer growth. It is clear from many studies that
phospholipid flip-flop in biogenic membranes occurs very rapidly, within
a period of a few minutes. These studies also reveal that phospholipid
translocation in biogenic membranes occurs bi-directionally,
independently of the phospholipid head group, via a facilitated
diffusion process in the absence of metabolic energy input, and that
this type of transport requires specific membrane proteins. These
translocators have been termed biogenic membrane flippases, and they
differ from metabolic energy-dependent transporters (ABC transporters
and MDR proteins). No biogenic membrane flippases have been
characterized. This review briefly discusses the importance of biogenic
membrane flippases, the various assay methods used for measuring the
rate of phospholipid flip-flop, and the progress that has been made
towards identifying these proteins.







{kind=link}
{kind=link}